
2.1 EL PEPTIDOGLUCANO: COMPOSICIÓN QUÍMICA Y ESTRUCTURA
2.1.1 COMPOSICIÓN QUÍMICA Y ESTRUCTURA BÁSICAS DEL PEPTIDOGLUCANO
2.2 RELACIONES ENTRE ESTRUCTURA Y FUNCIÓN EN EL PEPTIDOGLUCANO
2.3 OTROS COMPONENTES DE LA PARED CELULAR DE BACTERIAS GRAM-POSITIVAS: LA MATRIZ
2.4 PARED CELULAR DE LAS BACTERIAS ÁCIDO ALCOHOL RESISTENTES
2.5 LA PARED DE BACTERIAS GRAM-NEGATIVAS
2.5.1 LA MEMBRANA EXTERNA DE BACTERIAS GRAM-NEGATIVAS
2.5.1.1 COMPOSICIÓN QUÍMICA Y ESTRUCTURA DE LA MEMBRANA EXTERNA
2.5.1.1.2 Lipopolisacárido (LPS)
![]()
1 INTRODUCCIÓN
La mayor parte de los procariotas posee una pared celular (P.C.) rígida rodeando al protoplasto. Las excepciones son los micoplasmas (dentro del dominio Bacteria) y algunas arqueas, como Thermoplasma.
Al microscopio electrónico se puede observar como una capa en íntimo contacto con la membrana citoplásmica, con un espesor que oscila entre 10 y 80 nm (según especies) -frente a los 8 nm de la membrana celular- , y con una estructura más o menos compleja, según los tipos bacterianos.
a) Las paredes celulares más frecuentes en eubacterias siguen dos modelos alternativos que, como veremos comparten un componente común: paredes de tipo Gram-positivo o de tipo Gram-negativo.
b) Unas pocas eubacterias (como las del gén. Planctomyces) poseen paredes a base de proteínas.
c) Las Arqueas poseen paredes diferentes a las de eubacterias y se pueden agrupar en diversos tipos.
El grueso de este capítulo está dedicado al estudio de las paredes Gram-positivas y Gram-negativas, con sus principales variantes, pero finalizaremos con una alusión a los principales modelos de paredes arqueanas.
Aunque el alumno realizará en prácticas de labororatorio la tinción de Gram, vamos a explicarla aquí brevemente. La tinción de Gram es la más importante entre las tinciones llamadas diferenciales (aquellas que no tiñen de la misma manera a todos los tipos de bacterias). Resumiendo, esta tinción consta de varios pasos:
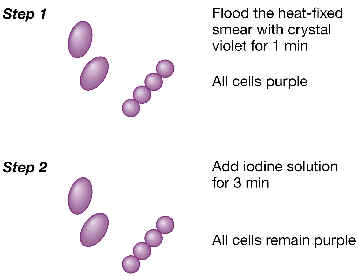 |
1. Imaginemos un frotis en porta de vidrio con una muestra de varios tipos de bacterias, que se han fijado por calor. En la primera fase, esta preparación de trata con un primer colorante llamado violeta cristal o violeta de genciana. 2. A continuación se añade una solución de lugol (yodo-ioduro), que actúa como “mordiente”, formando una laca relativamente resistente con el violeta. En este momento todas las bacterias están teñidas de color violeta. |
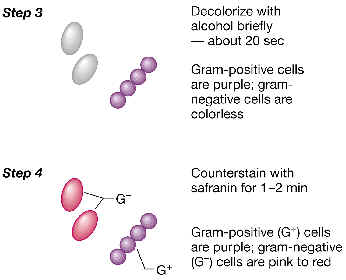 |
3. Ahora realizamos una descoloración diferencial con etanol o una mezcla de etanol y acetona. Lo que ocurre es que algunas bacterias (las que llamaremos Gram-negativas) pierden el color violeta (quedarían práctimante transparentes al microscopio), mientras que otras (las Gram-positivas) resisten el tratamiento, y retienen el colorante violeta. 4. Finalmente, tratamos el porta con un segundo colorante (colorante de contraste): se trata de un colorante de color rojo o rosa, como la fucsina o la safranina. Este colorante tiñe ahora a las bacterias previamente descoloradas por el etanol. Por lo tanto, el resultado en la observación al microscopio es que las bacterias Gram-positivas se ven de color violeta, y las Gram-negativas de color rosa o rojo. |
Como veremos enseguida, esta diferencia de comportamiento ante la tinción de Gram refleja el hecho de que ambos tipos de eubacterias poseen dos tipos estructuralmente diferentes de pared celular, aunque ambas posean en común la posesión de peptidoglucano. A continuación estudiaremos con cierto detalle la pared celular de eubacterias.
2 PAREDES DE LAS EUBACTERIAS
|
Consisten en un esqueleto macromolecular rígido, llamado peptidoglucano (= mucopéptido o mureína), que
|
 |
Comenzaremos abordando esa macrolécula tan peculiar llamada peptidoglucano, para después estudiar globalmente y por separado la pared de Gram-positivas y Gram-negativas, con algunas variantes que se pueden presentar.
2.1 EL PEPTIDOGLUCANO: COMPOSICIÓN QUÍMICA Y ESTRUCTURA
En las bacterias Gram-positivas el peptidoglucano representa el componente mayoritario de la pared celular (50-80% en peso), mientras que en Gram-negativas supone sólo del 1 al 10%.
2.1.1 COMPOSICIÓN QUÍMICA Y ESTRUCTURA BÁSICAS DEL PEPTIDOGLUCANO
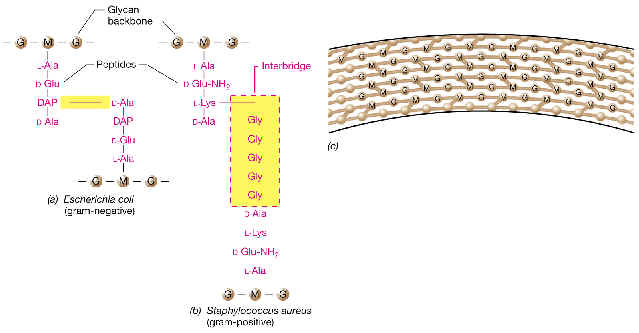
Está formado por repeticiones de una unidad disacarídica fundamental unida a su vez a un tetrapéptido. Distintas cadenas (formadas por el esqueleto de azúcares) se unen entre sí por determinados enlaces peptídicos entre tetrapétidos de cadenas diferentes. Veamos todo ello en su concreción química:
La unidad disacarídica repetitiva: consiste en N-acetilglucosamina (NAG) unida por enlace ß(1à4) a N-acetilmurámico (NAM). Obsérvese que el NAM es el 3-O-D-lactil-éter de la NAG (o sea, se deriva de unir el ácido D-láctico con el OH del C-3 de la NAG).
Las distintas unidades disacarídicas se van uniendo entre sí por enlaces ß(1à4) entre el NAM de una unidad y la NAG de la siguiente. Este enlace es susceptible a la rotura catalizada por el enzima lisozima. El número de repeticiones (n) puede oscilar entre 10 y 100.
La cadena tetrapeptídica: Desde el grupo carboxilo de cada ácido NAM, y mediante un enlace amido, se encuentra unido el tetrapéptido. Un tetrapéptido típico de muchas bacterias es:
L-alanina---D-glutámico---meso-diaminopimélico---D-alanina
Obsérvese la alternancia de aminoácidos D y L en el tetrapéptido.
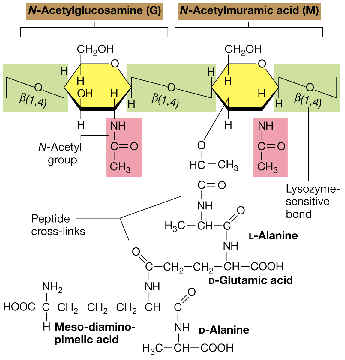
La estructura global: Las distintas cadenas polisacarídicas, con sus respectivos tetrapéptidos, se unen entre sí por medio de puentes o enlaces peptídicos, entre un aminoácido de una cadena (p. ej., el aminoácido nº3, como el meso-DAP del ejemplo) y otro aminoácido de una cadena adyacente (la D-ala terminal). De este modo, la estructura global es una sola macromolécula gigante que envuelve al protoplasto, formando un sáculo rígido, a modo de tejido continuo, que tiene el volumen y la forma de la bacteria respectiva.
|
En bacterias Gram-negativas este sáculo está formado por una sola capa (o unas pocas) de cadenas de PG. |
|
|
En Gram-positivas existen varias capas (hay varios niveles de PG). |
A continuación describiremos por separado el PG de Gram-positivas y Gram-negativas, dando indicaciones de sus principales variantes.
2.1.2 EL PEPTIDOGLUCANO DE BACTERIAS GRAM-NEGATIVAS
En la mayor parte de Gram-negativas el peptidoglucano corresponde a la composición y estructura que acabamos de describir. Sin embargo, en las espiroquetas, el diaminoácido en posición 3, en vez de ser meso-DAP, está sustituido por la L-ornitina (que también es un diaminoácido).
El enlace entre cadenas polisacarídicas se realiza normalmente mediante unión peptídica directa entre el grupo carboxilo de la D-ala terminal y el grupo e-amino del meso-DAP. Ahora bien, en este enlace participan solamente el 50% de los tetrapéptidos. Los demás péptidos no participan en enlaces, y entre estos últimos se encuentran incluso dipéptidos y tripéptidos.
El resultado es una capa simple de PG (de 1 nm de espesor), a modo de malla floja, y con grandes poros (los “huecos” dejados por las zonas donde no hay enlace peptídicos). Ello explica el comportamiento de las bacterias Gram-negativas en la tinción de Gram: al añadir el alcohol, se produce una deshidratación que tiende a contraer la estructura del PG, pero los poros son grandes y por ellos sale el primer colorante (el violeta de genciana). El ulterior tratamiento de la preparación con el colorante de contraste (fuchsina o safranina) tiñe a estas bacterias de rojo.
2.1.3 EL PEPTIDOGLUCANO DE BACTERIAS GRAM-POSITIVAS
Es más variado que el de Gram-negativas, sobre todo en función de ciertas variantes en la composición del tetrapéptido y del tipo de enlaces entre los tetrapéptidos.
|
Variantes en composición del tetrapéptido:
|
|||||||||||||||||||||
|
Modalidades de uniones interpeptídicas:
|
Desde el punto de vista estructural, el PG de Gram-positivas se caracteriza por la existencia de múltiples capas, existiendo entrecruzamientos tanto entre cadenas adyacentes en el mismo nivel como entre niveles distintos. El resultado es una red tridimensional gruesa (hasta 50 capas en algunos Bacillus), y más compacta que en Gram-negativas. De todas formas, el grado de compacidad varía entre especies, y depende de:
|
nº de NAM que contengan tetrapétidos que participen en entrecruzamientos; |
|
|
longitud del puente peptídico |
Ello condiciona a su vez la intensidad de la gram-positividad en la tinción de Gram.
2.2 RELACIONES ENTRE ESTRUCTURA Y FUNCIÓN EN EL PEPTIDOGLUCANO
La arquitectura molecular del sáculo de mureína está aún sujeta a debate. Sin embargo, uno de los modelos recientes más aceptado se podría resumir de la siguiente manera:
|
Los resultados de difracción de rayos X parecen indicar que las unidades disacarídicas de las cadenas están giradas unas respecto de otras, formando una estructura helicoidal de orden 4 o 5. A resultas de ello, los péptidos protruyen alternativamente: hacia arriba, a la izquierda, hacia abajo, a la derecha, y vuelta a empezar, etc. Esta organización permite que una cadena de PG se pueda unir con cadenas cercanas de su mismo nivel, así como con cadenas por encima y por debajo de su nivel, formando un perfecto entramado tridimensional (al menos en las Gram-positivas). |
|
|
La orientación de las cadenas azucaradas en relación con la superficie celular aún está debatida. Parece que se disponen casi paralelas a la superficie celular, con una tendencia a una forma espiral por encima de la membrana citoplásmica. Si consideramos una bacteria de forma bacilar, esta espiral cerrada se dispone siguiendo el perímetro circular (y no siguiendo el eje longitudinal). Los grupos tetrapeptídicos salen perpendicularmente de los NAM, en sentido vertical hacia la membrana. Sin embargo, cuando dos tetrapétidos de un nivel se unen entre sí, forman un puente casi horizontal, formando ángulos de unos 90o repecto de los esqueletos carbonados, y siguiendo el eje longitudinal de la célula. |
Esta estructura confiere una serie de importantes propiedades:
1) Gran rigidez, que contrarresta las fuerzas osmóticas a que está sometido el protoplasto (aguanta presiones de unas 5 a 15 atmósferas). Esta rigidez depende de:
a) el grado de entrecruzamiento;
b) el hecho de que el enlace ß(1à 4) es muy compacto. La alternancia regular entre anillos piranósicos de NAG y de NAM genera uno de los polisacáridos más estables desde el punto de vista termodinámico, que recuerda en su “estilo” a la quitina y a la celulosa;
c) la alternancia en el tetrapéptido, de aminoácidos en configuraciones D y L supone una factor adicional que confiere aún más fuerza estructural, y además permite que todas las cadenas laterales de estos aminoácidos se dispongan hacia el mismo lado, facilitando la formación de puentes de H.
2) Pero, al mismo tiempo, la estructura permite una notable flexibilidad. Ello colabora, junto con su rigidez, a soportar variaciones amplias de la tensión osmótica del protoplasto.
3) Condiciona la forma celular. Aunque la química del PG, por sí misma, no determina la forma, es su disposición espacial la responsable principal de esta forma.
2.3 OTROS COMPONENTES DE LA PARED CELULAR DE BACTERIAS GRAM-POSITIVAS: LA MATRIZ
Como ya dijimos, el PG de las bacterias Gram-positivas se encuentra inmerso en una matriz, que puede representar hasta el 50% del peso de la pared celular, y que está constituida por largos polímeros denominados ácidos teicoicos, pudiendo existir también (o en su lugar) los ácidos teicurónicos y los lipoteicoicos. Estos componentes llegan a sobresalir de la superficie celular y suministran especificidad antigénica.
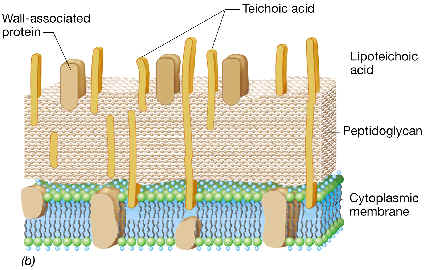
Ácidos teicoicos:
Están presentes en muchas bacterias Gram-positivas, pero no en todas. Son polímeros de hasta 30 unidades de glicerol-fosfato o ribitol-fosfato, unidas entre sí por enlaces fosfodiéster, en los que la mayoría de los grupos -OH están sustituidos por -H, azúcares, aminoazúcares o D-alanina.
Los ácidos teicoicos están unidos covalentemente al peptidoglucano, concretamente al -OH en posición 6 del NAM, a través de una unidad de enlace, variable según las especies. (Por ejemplo, en una especie de Micrococcus, el elemento de enlace consiste en {glicerol-P}3 --NAG-P).
Ácidos teicurónicos:
Ciertas bacterias Gram-positivas, cuando se someten a un régimen de limitación de fosfato son incapaces de sintetizar ácidos teicoicos, pero en su lugar producen ácidos teicurónicos. Los teicurónicos consisten en polímeros aniónicos formados por la alternancia de ácidos urónicos (que tienen grupos -COOH libres) y aminozúcares como la N-acetil-galactosamina.
Ácidos lipoteicoicos:
Están presentes en todas las bacterias Gram-positivas, aun en condiciones de carencia de fosfato. Se trata simplemente de ácidos glicerol-teicoicos que se encuentran unidos a la membrana citoplásmica, concretamente se unen por enlace fosfodiéster con glucolípidos de membrana, mientras que el otro extremo de la cadena queda expuesto al exterior.
En Streptococcus pyogenes las cadenas de lipoteicoicos se encuentran asociadas con la llamada proteína M, originando una microfibrillas que sobresalen notablemente hacia el exterior celular (observables a microscopio electrónico), y que facilitan la unión a las células de animales en que parasitan estas bacterias.
Funciones de los polímeros de la matriz:
|
Parece ser que su papel principal es suministrar una carga neta negativa a la pared celular, lo que permite captar cationes divalentes (p. ej., Mg++), que a su vez se necesitan para muchas actividades enzimáticas de la membrana citoplásmica o del espacio periplásmico, que participan de la morfogénesis y división de la pared celular. |
|
|
Como ya dijimos, los ácidos teicoicos y teicurónicos son buenos antígenos. Cuando no están cubiertos por estructuras más externas (como cápsulas), constituyen el antígeno somático O de las bacterias Gram-positivas. |
|
|
Finalmente, en algunas bacterias, sumistran, junto con el PG, receptores específicos para la adsorción de ciertos bacteriófagos. |
2.4 PARED CELULAR DE LAS BACTERIAS ÁCIDO ALCOHOL RESISTENTES
Determinadas bacterias Gram-positivas (corineformes, Nocardia y, en especial Mycobacterium) presentan una pared celular muy compleja, con abundancia de lípidos (algo excepcional entre las Gram-positivas).
Estas bacterias no se tiñen con los colorantes normales, pero una vez que se han teñido con fucsina (forzando mediante calentamiento de la preparación), tienen resistencia a decolorarse por una mezcla de ácido clorhídrico al 3% en etanol de 96o. Por ello se denominan como bacterias ácido-alcohol resistentes. Esta propiedad depende esencialmente de la presencia, en su pared celular de unos lípidos llamados ácidos micólicos.
Químicamente, esta pared celular consiste en un esqueleto formado por dos tipos de polímeros, unidos covalentemente entre sí:
|
un peptidoglucano especial (la diferencia más importante es que en vez de N-acetil murámico existe N-glucolil-murámico); |
|
|
un arabinogalactano de gran peso molecular. |
|
|
Ambos polímeros se encuentran enlazados a través de fosfodiéster entre una unidad de murámico y una de las arabinosas. Pero a su vez, este esqueleto se une covalentemente a los ácidos micólicos. |
Los ácidos micólicos son ß-hidroxiácidos grasos ramificados en a , cuya longitud de cadena es grande (desde C78 a C91 en Mycobacterium). Están unidos al esqueleto de la P.C. de forma uniforme, a través de enlaces con los -OH en 5 de las unidades de arabinosa.
Por lo tanto, el esqueleto de la P.C. de estas bacterias consiste en:
peptidoglucano---arabinogalactano---ácidos micólicos.
Pero aparte de este esqueleto complejo, la P.C. de las bacterias ácido-alcohol resistentes exhibe una variedad de lípidos:
1) Glucolípidos:
a) Micolatos de trehalosa: dos unidades de trehalosa unidas entre sí por enlace a (1à1´), y en donde los grupos 6 y 6´ están unidos con ácidos micólicos. Constituyen el llamado factor de crecimiento en cuerdas, debido a que son responsables de la agregación de los individuos bacterianos en forma de “cuerdas”.
b) Sulfolípidos de trehalosa: están localizados en la periferia de la P.C., y parecen ser impartantes factores de virulencia. En Mycobacterium tuberculosis (el bacilo de la tuberculosis) estos sulfolípidos de trehalosa funcionan como evasinas, es decir, facilitan el que la bacteria escape a la acción de los macrófagos inhibiendo la fusión del fagosoma con el lisosoma, lo cual puede explicar el hecho de que estosmicroorganismos tengan éxito como parásitos intracelulares.
c) Micósidos: Localizados en la periferia, consisten en la unión por enlace éster entre ácidos micólicos y azúcares (incluyendo ácidos urónicos, desoxiosas, aminozúcares, etc.).
2) Ceras: Unión de ácidos micólicos con ftioceroles (alcoholes ramificados de alto peso molecular: C30 - C34).
El alto contenido en lípidos confiere una serie de propiedades a estas bacterias (aparte de la ácido-alcohol resistencia ya citada):
|
aspecto y consistencia cérea de sus colonias; |
|
|
crecen formando grumos en medios líquidos; |
|
|
gran impermeabilidad de la P.C., que a su vez condiciona una gran resistencia a la desecación y gran resistencia a sustancias antibacterianas (detergentes, oxidantes, ácidos, bases, etc). |
2.5 LA PARED DE BACTERIAS GRAM-NEGATIVAS
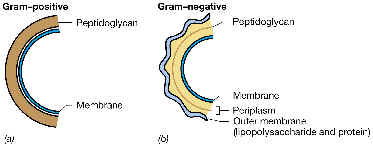
La pared de las bacterias Gram-negativas es estructuralmente más compleja que la de Gram-positivas, cosa que se puede comprobar al observar un corte transversal al microscopio electrónico: por encima de la membrana citoplásmica y hasta el exterior se pueden apreciar 5 capas, alternándose las claras (L2, L5) y las oscuras (más densas a los electrones: L1, L3, L4).
La capa densa L4 corresponde al peptidoglucano (ya estudiado), que se encuentra inmerso en la capa clara L5, que consiste en un espacio periplásmico, limitado entre la membrana citoplásmica y una membrana externa. La delgada capa de peptidoglucano no constituye más del 5-10% de la pared. A continuación estudiaremos la peculiar membrana externa de las bacterias Gram-negativas y el espacio periplásmico.
2.5.1 LA MEMBRANA EXTERNA DE BACTERIAS GRAM-NEGATIVAS
Se trata de una estructura de bicapa lipídica exclusiva de las bacterias Gram-negativas. Como otras bicapas lipídicas, consta de una doble capa de lípidos, junto con proteínas de matriz (estas últimas atravesando total o parcialmente la bicapa). Por supuesto, en la bicapa lipídica, los grupos polares quedan hacia afuera, mientras que los hidrófobos tienden al interior.
Ahora bien, la composición química y la disposición de los elementos de esta membrana externa son muy distintos a los de una membrana típica:
|
la bicapa es altamente asimétrica: |
|
|
en la capa externa existe un 60% de proteínas y un 40% de una macromolécula exclusiva de esta membrana externa: el lipopolisacárido (LPS); |
|
|
en la capa interna no hay LPS, existiendo fosfolípidos (FL), lipoproteínas (LPP) y otras proteínas. |
El conjunto es un mosaico fluido que permite el desplazamiento lateral de los fosfolípidos, del LPS, y de las proteínas, pero no de las lipoproteínas unidas covalentemente al peptidoglucano. Sin embargo, esta fluidez es menor que la de la membrana citoplásmica.
2.5.1.1 COMPOSICIÓN QUÍMICA Y ESTRUCTURA DE LA MEMBRANA EXTERNA
La membrana externa se encuentra unida con el peptidoglucano subyacente a través de distintos componentes y tipos de enlaces:
|
enlaces iónicos, mediados por cationes divalentes, entre distintas proteínas de la membrana externa y el PG; |
|
|
enlaces hidrófobos entre fosfolípidos y proteínas de la capa interior de la membrana externa con el PG; |
|
|
enlaces covalentes entre algunas moléculas de lipoproteína y el PG. |
Parece ser que la membrana externa está en contacto directo con la plasmática en numerosos puntos, denominados zonas de adhesión. Puede que se trate de regiones en las que produzca una auténtica fusión entre estos dos tipos de membrana, facilitando quizá el transporte de ciertas sustancias desde el exterior al interior celular.
Estudiaremos a continuación los componentes de la membrana externa, aludiendo a su composición química, estructura y funciones.
2.5.1.1.1 Fosfolípidos (FL)
Se localizan en la lámina interna de la m. ext. La composición en fosfolípidos es similar a la de la membrana citoplásmica, con un ligero enriquecimiento en fosfatidil-etanolamina.
2.5.1.1.2 Lipopolisacárido (LPS)
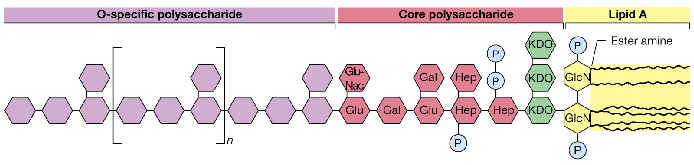
Se trata de una macromolécula exclusiva de la lámina externa de la membrana externa de bacterias Gram-negativas, responsable de muchas de las propiedades biológicas de estas bacterias. Se le conoce también con el nombre de endotoxina (toxina termoestable, no difusible). Se trata de un glucolípido complejo, que podemos considerar compuesto de tres regiones o dominios:
|
lípido A, que es la porción más proximal, y de carácter hidrofóbico; |
|
|
región intermedia, llamada oligosacárido medular; |
|
|
región distal (cadena lateral específica, polisacarídica) a base de repeticiones de unos pocos azúcares. Es de carácter hidrofílico y constituye el antígeno somático O de las 6bacterias Gram-negativas. |
i) El lípido A: esta región es prácticamente idéntica en todas las bacterias Gram-negativas. Consiste en un disacárido formado por dos unidades de glucosamina unidas por enlace ß(1à6), pero donde todos los grupos -OH (menos uno) y -NH2 están sustituidos (unidos a otras moléculas): Obsérvese que
|
existen 5 (a veces 6) ácidos grasos, todos ellos saturados, con predominio de ß-hidroximirístico (un ácido graso C14). |
|
|
El -OH original en 4´ está sustituido por arabinosamina-fosfato. |
|
|
El -OH en 1 está sustituido por fosforil-etanolamina (a veces pirofosforil-etanolamina). |
ii) El oligosacárido medular (también llamado corazón o núcleo): se une al lípido A a través del -OH en 3´. Se pueden considerar dos fracciones:
|
la fracción del núcleo interno, a base de dos tipos de azúcares exclusivos de Gram-negativas: 2-ceto-3-desoxioctónico (KDO) y L-glicero-D-manoheptosa (Hep). Alguna de las Hep y alguno de los KDO pueden a su vez estar unidos a fosforil-etanolamina (o pirofosforil-etanolamina). Esta región es muy rica en grupos cargados, especialmente con carga negativa (de los fosfatos y KDO). |
|
|
La fracción del núcleo externo está constituida a base de hexosas (glucosa, galactosa, NAG, y a veces algunas hexosas más raras). |
iii) Cadena lateral específica: polisacárido repetitivo, que se proyecta hacia el exterior celular, y que constituye el Ag somático O de bacterias Gram-negativas. Consiste en la repetición (hasta 40 veces) de unidades tri-, tetra- o pentasacarídicas (en estos dos últimos casos uno de los azúcares de cada repetición queda lateral respecto del esqueleto lineal que forman los demás).
De todas estas regiones del LPS la única indispensable para la viabilidad es el lípido A. Los mutantes incapaces de sintetizar las cadenas laterales o el oligosacárido medular dan colonias rugosas y están afectados en distintas propiedades biológicas, pero pueden sobrevivir.
PAPELES Y FUNCIONES DEL LPS
1. Papel estructural: el LPS es el componente esencial de la membrana externa. La porción hidrofóbica (las cadenas de ácidos grasos del lípido A) se proyectan hacia el interior de esta membrana. Precisamente es la estructura del lípido A la principal responsable de la menor fluidez de dicha membrana, y por lo tanto de la mayor resistencia física. (Obsérvese que, mientras cualquier fosfolípido tiene dos cadenas de ácido graso, el lípido A posee 5 o 6, todas ellas unidas al mismo disacárido, generando una molécula más “masiva.”).
2. A su vez, la propiedad anterior hace que sea menos soluble a detergentes y más resistente a disolventes orgánicos.
3. Es menos permeable a muchas moléculas hidrofóbicas, incluyendo antibióticos, debido a las largas cadenas laterales hidrofílicas.
4. Se une a cationes divalentes (como Mg++ o Zn++), lo que contribuye a la mayor estabilidad de la membrana externa. Esta presencia de cationes suministra un ambiente adecuado para muchas funciones de la P.C. (Si añadimos un agente quelante como el EDTA, o eliminamos el Mg++ y lo sustituimos por Ca++, se produce la desorganización de la membrana externa).
5. Como ya dijimos, el LPS constituye la endotoxina de las bacterias Gram-negativas. La función como endotoxina se debe a la región del lípido A. Sus propiedades como endotoxina están en el origen de muchos síntomas patológicos propiciados por patógenos Gram-negativos:
a. pirogenicidad (inducción de fiebre)
b. hipotensión
c. en casos graves, choque letal, por fallo cardíaco
d. actividad necrótica de tejidos.
6. Pero igualmente, tiene efectos beneficiosos: estimula una serie de mecanismos defensivos del hospedador, incluyendo la activación del complemento, que puede ocasionar la lisis de la bacteria, mejora las propiedades de los fagocitos, etc. Es decir, a pesar del nombre de endotoxina, el LPS no es intrínsecamente tóxico, sino que su efecto depende de la respuesta del hospedador. El macrófago es la célula del hospedador principal responsable de la mediación de los efectos del LPS, tanto los positivos como los negativos. El macrófago posee receptores de membrana para detectar el LPS bacteriano, y en respuesta a él, libera una serie de moléculas mediadoras (citoquinas) que actúan a su vez sobre diversas partes del sistema inmunitario. De hecho, los efectos negativos se deben a comportamientos incontrolados desencadenados en el propio sistema inmunitario.
7. El LPS, y concretamente las cadenas laterales constituyen el antígeno somático O, cuya especificidad viene determinada por la secuencia repetitiva de azúcares. Esta porción condiciona la virulencia de las bacterias Gram-negativas patógenas, por lo que debe de ser esencial en la interacción hospedador-parásito.
2.5.1.1.3 La lipoproteína (LPP, lipoproteína de Braun)
Su porción polipeptídica es una pequeña proteína (7.2 kDa) muy abundante en la membrana externa, y es la responsable de la unión covalente entre ésta y el peptidoglucano. La proteína tiene una configuración mayoritaria en a-hélice, que atraviesa el espacio periplásmico, y parece que se agrega formando trímeros. Una de las LPP del trímero (por término medio) se une covalentemente con el peptidoglucano.
El aminoácido N-terminal es una cisteína cuyo grupo sulfhidrilo está unido por enlace tioéter a un diglicérido, y cuyo grupo amino se une por enlace amido con un ácido graso (p. ej., palmítico). De este modo, la porción N-terminal de la LPP está embebida en la lámina interna de la membrana externa.
El aminoácido C-terminal es una lisina. Una de cada tres moléculas de LPP usa esta Lys para establecer un enlace peptídico entre su propio grupo -NH2 libre y el -COOH libre del meso-DAP del PG. Por término medio, por cada diez unidades disacarídicas del PG, existe un enlace covalente con la LPP.
La principal (y probablemente única) función de la lipopproteína es meramente estructural: estabilizar el complejo entre peptidoglucano y membrana externa. Esta unión es tan fuerte que permite aislar la membrana externa y el peptidoglucano como una unidad.
2.5.1.1.4 Proteínas de la membrana externa
Están intercaladas en esta membrana, participando en la estabilización de la arquitectura tridimensional, interaccionando unas con otras y con los lípidos. Entre ellas, las más importantes son las porinas.
Las porinas son proteínas de unos 35 kDa, que se agregan formando trímeros con canales interiores, y que atraviesan la membrana de parte a parte. Su función es permitir el paso de sustancias a través de dichos canales interiores, siempre que su peso molecular sea compatible con el tamaño de los canales (suelen ser moléculas entre 500 y 700 dalton). En las enterobacterias, las porinas colaboran en la protección contra las sales biliares que existen en el ecosistema intestinal donde pasan parte de su vida.
Existen otras proteínas minoritarias parecidas a porinas, que actúan como canales específicos que permiten el paso de ciertas moléculas: vitamina B12, quelatos de Fe, nucleósidos, maltodextrinas, etc. Algunas de ellas sirven simultáneamente como receptores de fagos.
2.5.1.2 PAPELES Y FUNCIONES DE LA MEMBRANA EXTERNA
1) Actúa como tamiz molecular, que permite la difusión únicamente de moléculas relativamente pequeñas. Esto supone una protección frente a muchos agentes antibacterianos: colorantes, ácidos biliares, antibióticos, enzimas (p. ej., la lisozima, que podría alcanzar y atacar al PG). Recuérdese que las porinas sólo permiten el paso de sustancias hidrofílicas por debajo del tamaño especificado por el diámetro de los canales.
2) Condiciona propiedades de superficie:
a) grado de humedad (humectabilidad)
b) adhesividad
c) carga eléctrica.
3) Es la estructura donde se fijan los componentes del complemento (sistema defensivo de los animales superiores que conduce a la inserción, en la membrana externa, de una serie de proteínas llamadas complejo de ataque a la membrana, que agujerea dicha membrana y ocasiona la lisis de la bacteria).
4) Ciertas proteínas y cadenas laterales del LPS pueden ser lugares de adsorción (receptores específicos) de fagos y bacteriocinas.
5) Punto de anclaje del anillo externo (“L”) del corpúsculo basal de los flagelos.
Entre la membrana externa y la membrana citoplásmica existe un compartimento acuoso bañando al peptidoglucano, denominado periplasma o espacio periplásmico. El volumen de este compartimento puede llegar a representar un 20-40% del volumen celular total. El contenido del periplasma (el “gel periplásmico”) incluye:
|
RNasa y fosfatasa, que digieren moléculas que por sí mismas no pueden pasar al citoplasma. |
|
|
Penicilinasa: degrada penicilina, evitando destrucción de PG |
|
|
proteínas de transporte de nutrientes (p. ej., de maltosa) |
|
|
proteínas de transporte de nutrientes (p. ej., de maltosa) |
|
|
proteínas de unión a estímulos químicos. |
|
|
En bacterias desnitrificantes y quimiolitoautotrofas, existen proteínas implicadas en el transporte de electrones. |
El periplasma cumple una función de osmorregulación: El periplasma es una solución densa, con alta concentración de macromoléculas, y que participa en la regulación de la osmolaridad celular frente a la tonicidad del medio exterior. Para ello, existe en este espacio periplásmico un oligosacárido derivado de la membrana citoplásmica (ODM, o según sus iniciales inglesas, MDO), a base de 10 unidades de ß-D-glucosa (unidas entre sí por enlaces 1à2) con sustituyentes ácidos.
|
En medios de alta osmolaridad (por ejemplo, en los fluidos corporales), disminuye la concentración del oligosacárido. |
|
|
En ambientes de baja osmolaridad (p. ej., aguas fecales), aumenta mucho la concentración de dicha molécula. De este modo, la presión de turgor del protoplasto se transmite contra el peptidoglucano, que como sabemos ya, es la estructura de la pared celular. que aguanta las variaciones de presión osmótica. |
3 PAREDES DE LAS ARQUEAS
Aunque las arqueas pueden comportarse como Gram-positivas o como Gram-negativas, no se suele aludir a esto en este dominio de procariotas, ya que sus paredes tienen poco que ver con las de eubacterias. Exceptuando el género Thermoplasma, carente de pared celular, las demás arqueas poseen, por encima de la membrana citoplásmica, algún tipo de estructura con funciones de pared celular.
|
En muchos casos las funciones de pared celular son ejercidas simplemente por una capa S paracristalina (véase tema 4), a base de disposición regular de subunidades idénticas de una proteína o glucoproteína (p. ej., Methanococcus, Methanogenium). Recuerde que en ciertas arqueas de ambientes extremos, esta capa S está estabilizada por factores de esos ambientes: En el halófilo obligado Halobacterium las subunidades de glucoproteína se estabilizan por altas concentraciones de ión Na+. En el termoacidófilo Sulfolobus la estabilidad la confieren los bajísimos pH. |
|
|
En Methanospirillum y en Methanothrix varias células, cada una con su capa S, se encuentran englobadas por una vaina común, a base de proteínas y carbohidratos, con una estructura a base de anillos paralelos. |
|
|
En Methanosarcina y Halococcus la capa S se encuentra rodeada de metanocondroitina, un polímero a base de N-acetil-galactosamina, glucurónico, glucosa y manosa. (Esta metanocondroitina es similar al condroitín sulfato presente en tejido conjuntivo de animales). |
|
|
Finalmente, los miembros del orden Methanobacteriales poseen una pared de pseudomureína, un extraño peptidoglucano no basado en NAM. Consiste en un esqueleto de unidades repetitivas de NAG unidas por enlace ß(1à3) con N-acetil-talosaminourónico (NAT, un azúcar exclusivo de estos organismos). El grupo -NH2 del NAT va unido a su vez con un tetrapéptido, pero en éste sólo participan aminoácidos de la serie L. Al igual que en la mureína de las eubacterias, las diversas cadenas se unen entre sí por enlaces peptídicos entre el aminoácido terminal (4) de un tetrapéptido y el diaminoácido (3) de otra cadena.
|
MAS
![]()
Índice de sesiones de diapositivas
Sitio web para la consulta de Microbiología general
Licenciatura de Biología, Universidad de Granada
esta es como la mesita, o sea dejanos tu sugerencias  franja morada veterinaria unne franja morada veterinaria unnePromocionar tu página también //
|
 algo de musiquita pa alegrar el día |




